
L’humanisme est-il contre-nature ?

CLAUDE BRAUN
Administrateur et éditeur en chef du "Québec humaniste"
Claude Braun a été professeur de neurosciences cognitives à l'UQAM de nombreuses années. Retraité depuis peu, Il a publié nombres de documents de recherches sur le sujet. Il a été également éditeur du "Québec laïque" et est depuis quelques années l'éditeur en chef de notre revue "Québec humaniste" Il a également publié "Québec Athée" en 2010. Téléchargeable gratuitement en utilisant ce lien avec les compliments de l'auteur.
Si on s’en était tenu à E.O. Wilson, jusqu’à récemment, l’humanisme, tel qu’incarné dans les principes officiels de l’Association humaniste du Québec, eut été jugé tout-à-fait contre-nature. Ce grand entomologiste évolutionniste, spécialiste des insectes sociaux, fut pendant 38 ans, protagoniste enthousiaste de la théorie de la parentèle comme explication complète et suffisante de l’altruisme dans le monde animal, humain compris. Cette élégante et séduisante théorie, formulée par William Donald Hamilton en 1964, stipule que la sélection naturelle de l’altruisme est une fonction directe et exclusive du degré de partage génétique entre le récipiendaire et le bénéficiaire de l’acte altruiste. Ce cadre d’analyse est centré sur le gène et l’organisme individuels. La sélection naturelle opèrerait exclusivement à ces niveaux. Cependant, dans son dernier essai [The social conquest of earth], E.O. Wilson rejette soudainement et radicalement la théorie de la parentèle comme explication suffisante de la sélection naturelle de la coopération animale et de l’altruisme humain [1]. Il propose à sa place, comme base biologique dominante de la coopération, et à fortiori de l’altruisme, la sélection de groupe. Ouvre-t-il vraiment, de cette façon, la porte à une harmonisation de la théorie de l’évolution et de l’humanisme ? Pas du tout ! Sous la plume de E.O. Wilson l’humain devient capable d’altruisme de très haut niveau, bien au-delà des limites de la parentèle, mais dans son fondement biologique il ne dépasse pas l’attachement au groupe : tribalisme, religion guerrière, superstition et rituel, sacrifice humain, racisme, xénophobie, patriotisme, militarisme, endoctrinement, etc. Wilson propose d’ailleurs de transcender notre biologie « tribaliste » et invoque désespérément la nécessité d’organiser notre monde culturellement, au-delà de notre biologie et même contre elle.
La théorie darwinienne de la sélection naturelle fut « déécologisée » par la découverte de l’ADN
 La « synthèse moderne » de la théorie de l’évolution depuis la découverte de l’ADN (c.f. Morgan et Weissman autrefois, Dawkins et Pinker aujourd’hui) a consisté beaucoup à voir le génome d’un individu comme étant l’unité essentielle de la sélection naturelle. Dans cette conception, c’est le génome de l’individu qui déterminera son développement et son comportement reproducteur. Que ce comportement soit social ou pas n’a aucune importance. Seule la quantité de progéniture apte elle-même à se reproduire compte. Cette conception est centrée sur le fait que la stabilisation de l’aptitude en question relève d’un ou plusieurs gènes de cet individu qui ne sont transmissibles que par ce même individu. Selon cette vision de la sélection naturelle, chaque gène, et par extension chaque génome, de chaque organisme individuel, a pour seule et unique fonction, du point de L’humanisme est-il contre-nature ? Claude Braun Si on s’en était tenu à E.O. Wilson, jusqu’à récemment, l’humanisme, tel qu’incarné dans les principes officiels de l’Association humaniste du Québec, eut été jugé tout-à-fait contre-nature. Ce grand entomologiste évolutionniste, spécialiste des insectes sociaux, fut pendant 38 ans, protagoniste enthousiaste de la théorie de la parentèle comme explication complète et suffisante de l’altruisme dans le monde animal, humain compris. Cette élégante et séduisante théorie, formulée par William Donald Hamilton en 1964, stipule que la sélection naturelle de l’altruisme est une fonction directe et exclusive du degré de partage génétique entre le récipiendaire et le bénéficiaire de l’acte altruiste. Ce cadre d’analyse est centré sur le gène et l’organisme individuels. La sélection naturelle opèrerait exclusivement à ces niveaux. Cependant, dans son dernier essai [The social conquest of earth], E.O. Wilson rejette soudainement et radicalement la théorie de la parentèle comme explication suffisante de la sélection naturelle de la coopération animale et de l’altruisme humain [1]. Il propose à sa place, comme base biologique dominante de la coopération, et à fortiori de l’altruisme, la sélection de groupe. Ouvre-t-il vraiment, de cette façon, la porte à une harmonisation de la théorie de l’évolution et de l’humanisme ? Pas du tout ! Sous la plume de E.O. Wilson l’humain devient capable d’altruisme de très haut niveau, bien au-delà des limites de la parentèle, mais dans son fondement biologique il ne dépasse pas l’attachement au groupe : tribalisme, religion guerrière, superstition et rituel, sacrifice humain, racisme, xénophobie, patriotisme, militarisme, endoctrinement, etc. Wilson propose d’ailleurs de transcender notre biologie « tribaliste » et invoque désespérément la nécessité d’organiser notre monde culturellement, au-delà de notre biologie et même contre elle. Vue de la sélection naturelle, d’assurer la survie d’autant de progéniture que possible. Plus précisément, comme le martèle avec insistance Dawkins, « l’ADN serait la matière la plus répliquante connue de l’univers ». Ainsi, tout comportement d’apparence altruiste (comportement favorisant la survie d’un autre aux dépens de sa propre survie) ne sera explicable que par l’une ou l’autre variante de la théorie de la parentèle. Pour simplifier, l’altruisme serait en fait un égoïsme génétique, selon la théorie de la parentèle, et le vecteur de la sélection naturelle resterait très exactement le gène et rien d’autre. Par extension, comporte mentalement, l’altruisme ne pourrait dès lors être conceptualisé biologiquement que comme échange réciproque de faveurs, et non comme « véritable » altruisme tel que l’entend le langage populaire. Par ailleurs, la théorie « individualiste » de la sélection naturelle postule que chaque organisme est en compétition avec tous les autres organismes pour survivre et se reproduire autant qu’il le peut. Sur le long terme ne seront donc sélectionnés que des gènes pouvant se répandre sur la planète terre autant que possible, tout simplement. Bref, la sélection naturelle est conçue ici comme opérant strictement à l’intérieur du théâtre de l’individu face au monde entier. Finalement, la théorie « individualiste » de la sélection naturelle tend à prendre une perspective « bottom-up » ou moléculaire, plutôt que « top-down» ou écologique. Bien entendu tout biologiste aujourd’hui admet que le gène réagit à l’environnement. Le gène « canonique » ou gène de « structure » détecte un signal dans la cellule qui le met en action pour produire de l’ARN qui synthétisera une protéine. Mais la subtilité des signaux potentiellement efficaces est généralement peu considérée par les tenants de l’inexistence de la sélection de groupe (Dawkins, Pinker, etc.). Le signal auquel un gène sera sensible n’est conçu comme pouvant être d’origine culturelle qu’avec réticence : on s’en tient à la mécanique moléculaire et cellulaire comme théâtre de détermination biologique. On insiste sur la mutation fortuite comme unique véritable cause de l’apparition de comportements sociaux dont l’avantage adaptatif apparaît dès lors comme « gratuit ».
La « synthèse moderne » de la théorie de l’évolution depuis la découverte de l’ADN (c.f. Morgan et Weissman autrefois, Dawkins et Pinker aujourd’hui) a consisté beaucoup à voir le génome d’un individu comme étant l’unité essentielle de la sélection naturelle. Dans cette conception, c’est le génome de l’individu qui déterminera son développement et son comportement reproducteur. Que ce comportement soit social ou pas n’a aucune importance. Seule la quantité de progéniture apte elle-même à se reproduire compte. Cette conception est centrée sur le fait que la stabilisation de l’aptitude en question relève d’un ou plusieurs gènes de cet individu qui ne sont transmissibles que par ce même individu. Selon cette vision de la sélection naturelle, chaque gène, et par extension chaque génome, de chaque organisme individuel, a pour seule et unique fonction, du point de L’humanisme est-il contre-nature ? Claude Braun Si on s’en était tenu à E.O. Wilson, jusqu’à récemment, l’humanisme, tel qu’incarné dans les principes officiels de l’Association humaniste du Québec, eut été jugé tout-à-fait contre-nature. Ce grand entomologiste évolutionniste, spécialiste des insectes sociaux, fut pendant 38 ans, protagoniste enthousiaste de la théorie de la parentèle comme explication complète et suffisante de l’altruisme dans le monde animal, humain compris. Cette élégante et séduisante théorie, formulée par William Donald Hamilton en 1964, stipule que la sélection naturelle de l’altruisme est une fonction directe et exclusive du degré de partage génétique entre le récipiendaire et le bénéficiaire de l’acte altruiste. Ce cadre d’analyse est centré sur le gène et l’organisme individuels. La sélection naturelle opèrerait exclusivement à ces niveaux. Cependant, dans son dernier essai [The social conquest of earth], E.O. Wilson rejette soudainement et radicalement la théorie de la parentèle comme explication suffisante de la sélection naturelle de la coopération animale et de l’altruisme humain [1]. Il propose à sa place, comme base biologique dominante de la coopération, et à fortiori de l’altruisme, la sélection de groupe. Ouvre-t-il vraiment, de cette façon, la porte à une harmonisation de la théorie de l’évolution et de l’humanisme ? Pas du tout ! Sous la plume de E.O. Wilson l’humain devient capable d’altruisme de très haut niveau, bien au-delà des limites de la parentèle, mais dans son fondement biologique il ne dépasse pas l’attachement au groupe : tribalisme, religion guerrière, superstition et rituel, sacrifice humain, racisme, xénophobie, patriotisme, militarisme, endoctrinement, etc. Wilson propose d’ailleurs de transcender notre biologie « tribaliste » et invoque désespérément la nécessité d’organiser notre monde culturellement, au-delà de notre biologie et même contre elle. Vue de la sélection naturelle, d’assurer la survie d’autant de progéniture que possible. Plus précisément, comme le martèle avec insistance Dawkins, « l’ADN serait la matière la plus répliquante connue de l’univers ». Ainsi, tout comportement d’apparence altruiste (comportement favorisant la survie d’un autre aux dépens de sa propre survie) ne sera explicable que par l’une ou l’autre variante de la théorie de la parentèle. Pour simplifier, l’altruisme serait en fait un égoïsme génétique, selon la théorie de la parentèle, et le vecteur de la sélection naturelle resterait très exactement le gène et rien d’autre. Par extension, comporte mentalement, l’altruisme ne pourrait dès lors être conceptualisé biologiquement que comme échange réciproque de faveurs, et non comme « véritable » altruisme tel que l’entend le langage populaire. Par ailleurs, la théorie « individualiste » de la sélection naturelle postule que chaque organisme est en compétition avec tous les autres organismes pour survivre et se reproduire autant qu’il le peut. Sur le long terme ne seront donc sélectionnés que des gènes pouvant se répandre sur la planète terre autant que possible, tout simplement. Bref, la sélection naturelle est conçue ici comme opérant strictement à l’intérieur du théâtre de l’individu face au monde entier. Finalement, la théorie « individualiste » de la sélection naturelle tend à prendre une perspective « bottom-up » ou moléculaire, plutôt que « top-down» ou écologique. Bien entendu tout biologiste aujourd’hui admet que le gène réagit à l’environnement. Le gène « canonique » ou gène de « structure » détecte un signal dans la cellule qui le met en action pour produire de l’ARN qui synthétisera une protéine. Mais la subtilité des signaux potentiellement efficaces est généralement peu considérée par les tenants de l’inexistence de la sélection de groupe (Dawkins, Pinker, etc.). Le signal auquel un gène sera sensible n’est conçu comme pouvant être d’origine culturelle qu’avec réticence : on s’en tient à la mécanique moléculaire et cellulaire comme théâtre de détermination biologique. On insiste sur la mutation fortuite comme unique véritable cause de l’apparition de comportements sociaux dont l’avantage adaptatif apparaît dès lors comme « gratuit ».
Il faut maintenant déboulonner de telles notions « néodarwiniennes » erronées comme celles qui suivent:
- Les gènes qui comptent vraiment sont ceux, les GÈNES DE STRUCTURE, qui codent pour une protéine. C’est la « production » de la protéine qui alimente le développement ainsi que le comportement de l’organisme ;
- Le moteur de la sélection de nouvelles espèces ou de nouveaux phénotypes est la MUTATION SPONTANÉE d’un gène de structure à la fois ;
- L’ADN est la structure la plus « répliquante » de l’univers ;
- Le gène produira une protéine sous l’exclusive détermination de l’acide désoxyribonucléique (ADN), les autres facteurs d’activation/désactivation n’étant que des modulateurs secondaires ;
- Un organisme vit seul dans sa peau et ne porte que son propre génome. Le phénotype est l’ORGANISME INDIVIDUEL, c’està-dire l’enveloppe externe du génotype ;
- Le succès évolutif d’une espèce se mesure à la QUANTITÉ d’ADN et de CYTOPLASME de cette espèce sur la planète. Ce concept fut longtemps capté dans l’expression « biomasse », souvent utilisée pour qualifier le succès évolutif d’espèces vivantes ;
La nouvelle synthèse écologiste de la sélection naturelle
Ces six postulats sont faux. Voici comment ils peuvent être reformulés correctement, en accord avec les connaissances actuelles en biologie.
1.Le génome d’une espèce animale est organisé dans une hiérarchie complexe d’influences modulatrices pouvant modifier, avec une petite mutation, le développement de systèmes corporels complexes et diversifiés ainsi que de générer des comportements complexes. Dans des organismes complexes comme les animaux sociaux la hiérarchie de contrôle des gènes de structure est beaucoup plus importante (par exemple du point de vue de leur masse) que les gènes de structure eux-mêmes. Après tout, les codons humains ne codent que pour 20 acides aminés (constitutifs des protéines) alors que des milliers de métabolites régulent l’action des gènes. C’est le jeu global des inactivations et activations stratégiques des gènes de structure qui détermine le développement et le comportement. Les SIGNAUX NON GÉNÉTIQUES déterminent davantage les signaux génétiques que l’inverse ;
- Le moteur de la sélection de nouvelles espèces ou de nouveaux phénotypes comportementaux est l’entrejeu intense et dynamique de variations écologiques et de variations génétiques proximales dans l’espace et le temps que l’on dénomme aujourd’hui l’ÉPIGENÈSE. L’environnement variable engendre et maintien des gènes de « réserve », récessifs, par exemple, qui déterminent des phénotypes minoritaires dans les populations, ou requérant des signaux déclencheurs rares, qui servent les populations et non les individus, parce que ces gènes « remémorent et anticipent » des circonstances environnementales rares;
- On sait depuis peu que L’ADN est très VOLATIL. Il mue spontanément à grande échelle et son expression (copie en ARN) est modulée par une multitude de métabolites. Par ailleurs, de nombreuses structures non vivantes sont beaucoup plus stables et « répliquantes » que l’ADN. Finalement, la nature a tout fait pour accroitre le « brassage » des gènes, par exemple en instaurant presque partout la reproduction sexuée;
- L’environnement détermine le moment et l’endroit où sera produit une protéine, davantage que l’ADN, car les gènes ne sont activés ou désactivés que par des métabolites-signaux. À mesure que l’organisme se développe, toujours en interaction constante avec un environnement beaucoup plus complexe que lui, des milliards de différentes influences environnementales impactent le génome et le régulent. La structure la plus répliquante de l’univers n’est pas le gène. Du point de vue de la vie, la structure la plus répliquante est le SYSTÈME PLANÉTAIRE lui-même: la terre n’a pas manqué une seule rotation autour du soleil depuis quatre milliards d’années;
- Un organisme vit en communauté avec d’autres organismes dans sa peau et aussi dans son environnement immédiat (sa niche écologique) : il est SUPER-ORGANISME, et de même, une colonie d’animaux sociaux fonctionne, elle-aussi comme un super-organisme. À partir du moment où une espèce devient « co-habitante » (ex : nuée d’insectes vivant dans une même niche), ce sont les particularités sociales qui dominent la sélection naturelle ;
- Le succès évolutif d’une espèce se mesure à l’INTERVALLE DE TEMPS pendant lequel elle peut persévérer, dont l’élément clé est sa contribution à une niche écologique harmonieuse. Par exemple, il est rare qu’un virus ou qu’une bactérie tue son hôte de prédilection car si tel était le cas, il perdrait rapidement sa source alimentaire. Il ne tuera que des hôtes servant à le transporter vers son hôte de prédilection ;
La cellule est un super-organisme
 Chaque cellule du corps humain est unique. Elle est unique en grande partie en raison du voisinage des autres cellules, voisinage qui se modifie et se différencie dramatiquement au long du développement, et ensuite au long des échanges entre l’organisme et le reste du monde, incluant le vécu social. Les cellules environnantes s’échangent des signaux, ce qui canalise et individualise leur développement. La cellule individuelle est elle-même un « village ». On trouve à proximité de l’ADN toutes sortes de visiteurs, provenant d’autres « villages », qui modulent l’action de segments de cet ADN. Ces visiteurs proviennent de l’environnement moléculaire intra et extracellulaire et même extra-organismique. Ce village est capable de réparer des bris chromosomiques. Des « visiteurs » sont, à l’occasion, capables de se coller sur certaines parties de l’ADN, de façon assez tenace pour que leur modulation de l’activité génomique soit transmissible à la progéniture, cela sans que la structure de l’ADN ne soit modifiée, sans mutation de quelque gène que ce soit. L’effet de l’ADN est donc modifié par le vécu de la cellule. Le gène ne fonctionne pas comme un Dieu créateur. Il n’est qu’un membre d’un groupe à l’intérieur même de la cellule et il dépend du vécu global de ce groupe. Si le cytoplasme d’une cellule n’était pas un « village », tous les organismes ne seraient rien d’autre que des amalgames de clones d’ovules.
Chaque cellule du corps humain est unique. Elle est unique en grande partie en raison du voisinage des autres cellules, voisinage qui se modifie et se différencie dramatiquement au long du développement, et ensuite au long des échanges entre l’organisme et le reste du monde, incluant le vécu social. Les cellules environnantes s’échangent des signaux, ce qui canalise et individualise leur développement. La cellule individuelle est elle-même un « village ». On trouve à proximité de l’ADN toutes sortes de visiteurs, provenant d’autres « villages », qui modulent l’action de segments de cet ADN. Ces visiteurs proviennent de l’environnement moléculaire intra et extracellulaire et même extra-organismique. Ce village est capable de réparer des bris chromosomiques. Des « visiteurs » sont, à l’occasion, capables de se coller sur certaines parties de l’ADN, de façon assez tenace pour que leur modulation de l’activité génomique soit transmissible à la progéniture, cela sans que la structure de l’ADN ne soit modifiée, sans mutation de quelque gène que ce soit. L’effet de l’ADN est donc modifié par le vécu de la cellule. Le gène ne fonctionne pas comme un Dieu créateur. Il n’est qu’un membre d’un groupe à l’intérieur même de la cellule et il dépend du vécu global de ce groupe. Si le cytoplasme d’une cellule n’était pas un « village », tous les organismes ne seraient rien d’autre que des amalgames de clones d’ovules.
L’animal est un super-organisme
Chez l’animal, ce qui est sélectionné et ce qui se reproduit ce ne sont pas qu’UN génome d’UN organisme. Cela est une abstraction et une fausseté. Ce qui est sélectionné, et ce qui se reproduit est PLUSIEURS génomes de PLUSIEURS organismes. D’abord, c’est un couple qui se reproduit et non un individu, une première variante de sélection de « groupe ». Ensuite, de nombreuses plantes ne sont absolument pas viables autrement qu’amalgamées avec d’autres plantes et insectes avec lesquels elles vivent en symbiose. La sélection naturelle de tels organismes est une sélection de groupe. La symbiose est critique aussi pour les humains qui sont composés en fait de plusieurs génomes. Par exemple, 90% des cellules du corps humain sont des organismes étrangers (protistes), eux-mêmes porteurs de leurs propres génomes. Ces « parasites » peuvent être maléfiques, mais la plupart sont bénéfiques, absolument nécessaires pour la survie de l’organisme qui est en fait un multi organisme, multi-génomique. Un humain sans flore bactérienne n’est pas viable. Cette flore participe au développement de l’organisme de façon critique, à sa digestion, ainsi qu’à plusieurs autres aspects de son homéostasie dans plusieurs autres systèmes corporels, incluant le cerveau. Le génome humain lui-même est composé de vestiges de nombreux organismes très différents. Par exemple, environ 10% du génome humain consiste en vestiges de virus. De même, chaque cellule de notre corps comporte des milliers de mitochondries, petites usines de production énergétique, portant chacune son propre ADN. Leur ADN, lorsque mutant, est capable de détruire le cerveau, le squelette, la peau, les viscères. À l’inverse, leur ADN est probablement important dans le développement normal de ces mêmes systèmes corporels. Les mitochondries sont des vestiges évolutifs bactériens autrefois partiellement phagocytés.
Donc, qu’est-ce qui est transmis lorsqu’un humain se reproduit ? Il y a d’abord les gènes des parents, cela va de soi, 50% provenant de chaque parent. Mais il y a aussi les protistes de la mère, ainsi que l’ADN pseudo-bactérien (mitochondrie) de la mère. Finalement, il y a aussi une multitude de signaux moléculaires se rendant au génome provenant directement ou indirectement de l’environnement externe et de l’environnement interne et qui en régulent la fonction. La régularité transmissible n’est donc pas entièrement dépendante du génome parental. Elle est aussi tributaire d’autres génomes et d’autres organismes ainsi que de signaux critiques fournis par la partie non vivante de la niche écologique (ex : pression barométrique, humidité, température, variables qui se multiplient à l’infini : pensons par exemple à ce poisson, Odontesthes bonariensis, qui change de sexe selon la température de l’eau). Pour résumer, ce qui est sélectionné est un super-organisme ou multi-organisme, bref, un groupe.
La collectivité est un super-organisme
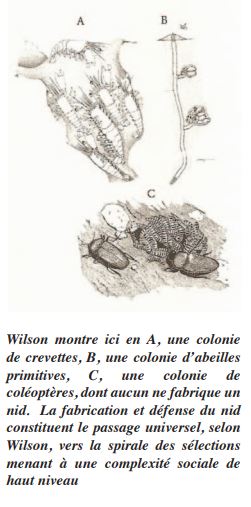
Les hyménoptères ainsi que toute autre espèce occupant un nid (ou une collectivité) à vie en grand nombre fonctionnent comme un super-organisme [1]. Lorsque la reine a un bébé abeillette, cette larve reçoit non seulement les gènes de ses deux parents (surtout de la mère), mais elle reçoit aussi de nombreux signaux biologiquement prégnants, même génétiquement déterminants, provenant entre autres de la présence et l’activité des autres larves, des abeilles adultes, etc. Qu’est-ce qui déterminera l’apparition d’une nouvelle espèce d’insectes sociaux à partir d’une espèce non sociale? Est-ce une mutation massive de nombreux gènes qui d’un seul coup créent un hyménoptère vivant à 40,000 dans un nid qu’il a lui-même construit ? Ou n’est-ce pas plutôt un nouvel environnement, accompagné de l’inhibition d’un gène de dispersion, suivi de petites mutations à débit rapide touchant délicatement des traits « sociaux » préadaptés, tel que le propose Wilson [1] ? Il est dépassé aujourd’hui de penser que la collectivité hypersociale (ex : hyménoptères) ne soit possible que chez des clones. Wilson montre comment des collectifs animaux hypersociaux peuvent exister avec un partage génétique beaucoup moindre que les hyménoptères (abeilles, guêpes, fourmis) … [1].
Un gène est typiquement positionné de façon à ne s’activer que sous l’influence d’un événement environnemental. Cet événement peut être extrêmement régulier, tel le passage des saisons, la présence d’eau ou d’oxygène. Cet événement émerge donc d’un environnement davantage RÉPLIQUANT que les génomes eux-mêmes. Ce qui fait que de considérer le seul gène comme agent de « réplication » est une idée fausse, une mauvaise abstraction. L’environnement social a lui aussi un impact physique, moléculaire, sur le corps, et cet impact se rend jusqu’aux gènes, qui lui sont sensibles et y réagissent. Le léchage du raton par la rate-mère est critique pour que s’effectuent des cascades développementales endocriniennes et cérébrales de la progéniture. Des signaux socio-environnementaux déterminent laquelle abeille deviendra une guerrière et laquelle une ouvrière. Certains de ces signaux ont pour effet de se coller des molécules stratégiquement sur certains brins d’ADN afin de le moduler dans leur action pour plusieurs générations à venir. De même, chez l’humain, la famine ou la maltraitance extrême entraînent des variations de méthylation aux environs de gènes spécifiques faisant en sorte que de nouveaux phénotypes (morbides pour la plupart, mais pas tous…) se développeront et seront transmis à la progéniture. La compétition entre individus d’un groupe, par exemple entre humains, pour sortir vainqueur(s) de la sélection naturelle existe, bien entendu. Mais, ces multi-organismes multi-génomiques, ces « individus », vivent en groupes qui peuvent se faire une compétition farouche, en tant que groupes, à l’intérieur d’une même « espèce ». Un groupe peut annihiler un autre groupe. Ici, ce qui est sélectionné n’est pas un foisonnement reproducteur, mais une occupation exclusive d’une niche particulière.
Base biologique de l’altruisme:
Un génotype à expression « capricieuse » Revenons à la question de l’altruisme. Supposons que des groupes particuliers d’humains aient « bénéficié » d’une combinaison de gènes et de culture garantissant une façon de s’organiser de façon « altruiste » sur une autre base que la parentèle, mettons sur la base de l’attachement à la famille, au village, à la ville ou à la nation… Appelons cela, à l’instar de Wilson, le fanatisme… On peut supposer qu’un cerveau particulièrement fabulateur puisse en être un élément clé. À ce moment-là il devient tout à fait imaginable que ce groupe particulier puisse éliminer tout compétiteur (race, ethnie, tribu, groupe socio-économique) de la même espèce ou d’une autre espèce. Selon Wilson, les analyses fines par modèles mathématiques montrent de plus en plus que la théorie de la parentèle est beaucoup trop grossière pour expliquer les dispositions altruistes versus égoïstes des animaux et des humains. La sélection naturelle opère aussi via la guerre ou n’importe quelle autre forme d’extermination ! Chez l’humain, la guerre ce n’est pas seulement l’agressivité, c’est aussi l’acceptation du risque de mourir pour le groupe (ou de ne pas se reproduire), une éventualité qu’aucune formule parentéliste ne peut expliquer. Le groupe devient ainsi le théâtre de la sélection naturelle, et l’individu en compétition avec tout conspécifique devient une figure de l’esprit, une fausse abstraction. Ceci laisse amplement de place pour que des gènes soient sélectionnés dans les individus formant un groupe non pas parce que ces gènes favorisent la survie de l’individu mais parce qu’ils favorisent la survie du groupe. Dit de façon plus précise, la sélection de groupe importe dans la mesure où la vie de groupe est déterminante quant aux chances du membre d’avoir une progéniture partageant ne serait-ce que très peu de ses gènes. Imaginons cela de façon concrète. Lequel aura plus d’enfants aptes à se reproduire ? Le bushman qui vit en tout petits groupes peu organisés ou le citoyen d’un empire guerrier triomphant ? Poser la question, c’est y répondre. Les bushmen d’Afrique se sont d’ailleurs récemment mis sous la protection de l’ONU contre l’extermination par les bantous de la part desquels ils craignent l’extermination…
Chez l’humain, il n’y a pas que les gènes qui changent de millénaire en millénaire. Les écologies et les cultures changent aussi. Les gènes comportent un point de bascule entre la réplication et la mutation. Il en va de même pour la niche écologique. Mais c’est à l’intersection de ces deux bascules que l’évolution des espèces se fait. Certaines cultures ou économies humaines ont tellement changé qu’elles ont induit des pressions sélectives radicales sur certains gènes. Un exemple spectaculaire est celui du groupe de gènes, autrefois inutile, permettant à l’adulte humain de développer les enzymes capables de cataboliser le lactose afin de le digérer. C’est l’évolution culturelle-économique, une invention de groupes d’humains, qui a conduit à la domestication animale et à la récupération du lait animal sur une base quotidienne, voire même indispensable. Partout où la consommation du lait est devenue prévalente, le groupe de gènes en question est devenu prévalent lui-aussi. Les « individus » humains incapables de digérer le lait n’ont pas survécu: hécatombe, sélection. Mais pas si naturelle que ça, et pas si individuelle que ça. Voilà une interaction culture-gènes où c’est le groupe qui est le théâtre de la sélection bien plus que ne l’est l’individu et où c’est la culture qui change la donne avec le génome qui suit. Wilson croit qu’il en est de même pour l’établissement d’espèces d’insectes sociaux : la mutation génétique est presqu’insignifiante tandis que le nouvel environnement est lourdement déterminant [1].
Il est inutile pour un organisme du règne animal de couvrir la planète de son ADN. S’il fait cela, il disparaît, faute de sources alimentaires. Dès l’émergence du règne animal, la sélection naturelle ne fonctionne pas par emballements mais plutôt par accommodements écologiques. Seuls de tels accommodements peuvent garantir la pérennité d’un animal, qu’il soit protozoaire ou humain. La stratégie de l’emballement, de l’invasion, de la reproduction tous azimuts, n’est concevable que dans un monde où débute la vie. À ce moment-là, un seul gêne peut tout changer en effet, et sa réplication peut « occuper » le monde entier. Par exemple, un gène qui permet une photosynthèse particulièrement efficace suffit à offrir la planète à l’organisme hôte sur un plateau d’argent. L’altruisme de l’organisme hôte n’a à s’appliquer qu’aux membres de l’unique même espèce peuplant désormais la planète. Mais dans un monde comme le nôtre, pour un organisme comme le nôtre, l’altruisme doit s’appliquer à tout ce qui est favorable à la vie de groupe : la nature « inanimée » en équilibre, les autres espèces, les membres génétiquement éloignés de notre espèce, etc.
Une définition précise de la sélection de groupe
Pour résumer, la détermination de la trame des adaptations passe, dans la phylogenèse, de la mutation modifiant les réponses à l’environnement non social, chez les animaux asociaux, à la mutation favorisant l’adaptation sociale chez les animaux sociaux. Qu’est-donc exactement que la sélection de groupe ? Wilson n’en formule pas une définition claire. Mais on comprend, en le lisant, que ce qui est bon pour le groupe peut être mauvais, dans certaines circonstances, pour certains individus, et que, donc, tout ne se joue pas en termes de mutations affectant la capacité procréatrice des individus. Je vais essayer de formuler dans mes propres termes, aussi clairement que possible, l’idée de sélection de groupes, incompatible avec la théorie de la parentèle.
La sélection de groupe consiste en sélection de « facteurs » biologiques qui engendreront des phénotypes gagnants pour la capacité procréatrice à long terme de la communauté et même possiblement perdants à court terme pour la capacité procréatrice des individus porteurs des dits gènes. Toutefois, pour que de tels « facteurs » soient auto-répliquant, on doit supposer qu’ils soient activés par des circonstances environnementales particulières (non universelles pour l’espèce), et qu’en dehors de ces circonstances, ces « facteurs » ont un effet nul ou favorable sur la capacité reproductrice de l’individu.
Cette idée ouvre la possibilité de comportements suicidaires, asexuels, auto-sacrificiels, ou simplement altruistes à l’extrême. Ces comportements ne peuvent devenir la norme universelle pour TOUS les membres de l’espèce: ils doivent être circonstanciels ou ne se manifester que dans des sous-groupes. On trouve une multitude de tels comportements et de tels sous-groupes dans le monde animal. L’abeille qui darde se suicide elle-même (mais la circonstance est exceptionnelle) … L’étalon sauvage typique est asexuel (seul l’étalon dominant a accès aux femelles dans la troupe) mais le mâle alpha est hyperfertile… On retient de ce dernier exemple qu’il suffit de bien peu d’individus non-altruistes dans une espèce pour que l’altruisme se répande à très grande échelle. L’étalon alpha, dans d’autres circonstances, deviendra l’étalon beta, auto-castré. L’abeille qui darde contribue, tout en mourant, à la survie de sa progéniture existante ainsi que de sa nuée au complet. Le geste altruiste ne viole donc pas l’esprit de la règle de Hamilton, mais la complexifie. Le geste altruiste relève de mécanismes de sélection déterminés non plus entièrement par le rapport entre le dardeur et le dardé, mais par le rapport entre le dardeur, le dardé et la nuée. De même, le geste altruiste ne relève pas que de gènes mais de toute sa biologie.
Humanisme et natures humaines
L’humanisme est-il contre-nature ? Oui bien sûr ! Mais de quel humanisme et de quelle nature parle-t-on ici ? Comme nous l’avons vu plus haut, la nature biologique de l’altruisme humain, aussi spectaculaire qu’en soient les expressions, est essentiellement limitée au « tribalisme ».
Pour ce qui est de l’humanisme dont il est question ici, c’est l’idée qui accorde à tout humain la même dignité, les mêmes droits fondamentaux, et, en toute sincérité, exactement la même valeur.
Ce sont deux élans complètement contradictoires et incompatibles. L’humanisme est la seule doctrine qui s’oppose directement aux divers tribalismes, ethnocentrismes, religions, en proposant un standard éthique encore plus élevé.
L’humanisme est la culmination d’une autre nature humaine, notre nature culturelle [2]. Parce que nous sommes des animaux sociaux, cette nature culturelle de l’humain est capable de transformer radicalement nos mœurs. De la brute solitaire bienveillante mais extrêmement dangereuse que fut le premier Homo sapiens sapiens, nous nous sommes hissés au statut de brutes désarmées et réprimées, vivant en très grands groupes, bêtes beaucoup plus bienveillantes, capables de violence extrême seulement par trébuchement, conditions extrêmes, ou échec des mécanismes culturels de transmission des valeurs [3]. Mais ce n’est pas par angélisme que cette évolution culturelle s’est faite chez Homo sapiens. Comme l’explique élégamment Jared Diamond [4], le rapport qu’entretient l’écologie humaine à la violence est important mais paradoxal : le chasseur-cueilleur est naturellement pacifique. Il n’a pas de système culturel ritualisé d’apaisement, d’interdiction du meurtre, etc. Il n’en a pas besoin car il n’y a aucun gain dans l’écologie chasseur-cueilleur à exploiter, dominer, tuer… Cependant, il tue sans vergogne et sans remord lors d’occasions inhabituelles où un étranger ou quelques étrangers entrent en conflit avec lui. À l’opposé, le cultivateur-pasteur vit dans une culture qui ritualise et absolutise de nombreuses règles de pacifisme. La religion occupe la quasi entièreté de cet espace rituel et culturel. Mais ces valeurs ne s’appliquent chez lui que pour la tribu. Cette même tribu valorise et ritualise les massacres humains à grande échelle, pourvu qu’il s’agisse d’autres tribus.
L’humanisme, aujourd’hui, et c’est nouveau, est une anticipation intellectuelle et rationnelle de la niche écologique à laquelle l’espèce humaine sera inévitablement confrontée. Le seul humanisme conséquent aujourd’hui, face à la menace environnementale ainsi que face à l’Armageddon nucléaire, est l’éco-humanisme pacifiste. Partout au monde, les communautés humaines deviennent (ou deviendront) multiraciales, les cultures, mondiales, les sources alimentaires interdépendantes. Et les peuples seront de plus en plus capables de s’annihiler militairement, mutuellement. Nous n’avons pas le choix d’être humanistes, voir même éco humanistes et pacifistes. Nous serons méta-super-organisme, tous ensemble, ou nous ne serons pas. D’ici peu, ce ne sera pas seulement la survie des générations futures qui sera en jeu, mais … celle de chacun de nous au courant de notre propre vie. Cela aussi est dit, en toute limpidité, par Wilson dans son dernier très bel ouvrage.
L’auto-désélection comme altruisme ultime
J’ajoute toutefois ma propre touche d’ironie. Le sacrifice ultime, l’altruisme ultime, du point de vue biologique, est de ne pas se reproduire. Le monde en est justement rendu au point où ce sacrifice biologique, ce geste le plus ultimement contrenature, est exactement celui que notre raison, notre culture, notre moralité, nous dictent de poser. La cause majeure de la dégradation de l’environnement, et du risque de déflagration d’armements nucléaires, est la surpopulation. Il faut, pour la première fois de notre existence en tant qu’espèce, que nous réduisions dramatiquement notre taux reproductif. Pour ajouter une couche à cette ironie, soulignons que c’est l’éthologiste V.C. Wynne-Edwards qui a introduit l’idée de sélection de groupe en 1962 en voyant des populations de lemmings se jeter collectivement de falaises directement à leur mort lors d’épisodes de surpopulation [5].
- Wilson, E.O. (2012). The social conquest of earth. New York : Liveright Publishing Company.
- Barrette, C. Communication personnelle. Voir aussi «Le miroir du monde», 2000. Montréal : MultiMondes.
- Pinker, S. (2011). The better angels of our nature: Why violence has declined. New York : Viking.
- Diamond, J. (1997). Guns, germs, and steel. New York : W. W. Norton.
- Wynne-Edwards, V.C. (1962). Animal dispersion in relation to social behaviour. Edinburgh: Oliver & Boyd.
0 commentaires